Análisis multivariado en la comparación morfométrica del híbrido (♂Colossoma macropomum X ♀Piaractus brachypomus) y sus parentales
Multivariate morphometric analysis in the comparation of the hybrid (♂Colossoma macropomum X ♀Piaractus brachypomus) and their progenitors
Danny Villegas1![]() , Manuel Emilio
Milla Pino2*
, Manuel Emilio
Milla Pino2*![]() , Otto Castillo González1
, Otto Castillo González1![]() , Karina Durant1
, Karina Durant1
1 Universidad Nacional Experimental de los Llanos Occidentales Ezequiel Zamora, Programa de Ciencias del Agro y del Mar, Guanare, Venezuela
2 Universidad Nacional Experimental del Yaracuy, Yaracuy, Venezuela
*Autor de Correspondencia, e-mail manuel.milla@unj.edu.pe
Recibido, 17/04/2020 Aceptado, 22/04/2020 DOI:10.25127/aps.20201.546

RESUMEN
Realiza un análisis morfométrico multivariado a través del método de componentes principales y el análisis discriminante entre el híbrido cachamoto y sus parentales; (Colossoma macropomum) macho y el morocoto (Piaractus brachypomus) hembra. El material utilizado en el ensayo se obtuvo de los estanques de la estación piscícola Papelón, en el estado Portuguesa (Venezuela), donde se estudiaron un total de 33 ejemplares (11 híbridos, 11 ejemplares de cachama negra y 11 de cachama blanca), a cada pez se le tomaron 32 medidas siguiendo el método de las cerchas o “trusses” con base en la distancia entre puntos homólogos. Los resultados del análisis discriminante evidenciaron diferencias significativas (p < 0,05) entre el híbrido y sus parentales, en relación a variables morfológicas tales como: base aleta adiposa, posterior aleta dorsal al origen aleta anal, posterior aleta anal a la caudal, ancho pectoral (cabeza), posterior aleta dorsal a la posterior aleta adiposa, base aleta anal, aleta pélvica al origen aleta anal, articular al origen aleta pectoral, posterior aleta adiposa a la posterior aleta anal.
Palabras clave: híbrido, parentales, morfometría, multivariado, cerchas, Colossoma macropomum, Piaractus brachypomus.
ABSTRACT
In this paper presents a multivariate morphometric analysis through principal components and discriminant analysis method between cachamoto hybrid and its progenitors; (Colossoma macropomum) male and morocoto (Piaractus brachypomus ) female. Material used in the trial was obtained from the Papelón station fish ponds in Portuguesa state country (Venezuela), 33 specimens where studied (11 hybrids, 11 morocotos, and 11 black pacu), on each fish 32 variables were measured according to trusses method based on distances between homologous point. The discriminant analysis results showed significant differences (p<0.05) between hybrid and its parents, in relation to morphological variables such as: base adipose fin, posterior dorsal fin to anal fin origin, posterior anal fin to caudal, pectoral width (head), posterior dorsal fin to posterior adipose fin, base anal fin, pelvic fin to anal fin origin, articulating pectoral fin origin, posterior adipose fin to anal fin.
Keywords: hybrid, parent, morphometry, multivariate and trusses, Colossoma macropomum, Piaractus brachypomus.
I. INTRODUCCIÓN
La cachama Colossoma macropomum es un pez omnívoro que se alimenta de muchas clases de flores, frutos y plancton; su crecimiento es rápido y alcanza buen tamaño; es poco exigente en cuanto a la calidad del agua y puede soportar bajas concentraciones de oxígeno disuelto. Según Orozco (1990) la cachama blanca Piaractus brachypomus, es nativa de las cuencas de los ríos Orinoco y Amazonas y es considerada como la especie de mayor potencial productivo y comercial en la piscicultura extensiva, semi-intensiva e intensiva de aguas cálidas continentales.
Así mismo, en Venezuela se ha logrado con mucho éxito la hibridación de las dos especies de cachamas Colossoma macropomum y Piaractus brachypomus (Heredia y González, 1986), al punto que el 80% de las cachamas cultivadas corresponden a esta (Campos, 2007). Al mismo tiempo la implementación del análisis morfométrico en algunas especies aporta conocimientos científicos que ayudan al mejoramiento genético. Cabe destacar que, a pesar de todo, aún no es suficiente el conocimiento que se tiene en cuanto a la adecuada selección de los híbridos, ya que es difícil para algunos productores identificar estos individuos.
En este sentido este proyecto destaca la necesidad de buscar características morfológicas que permitan a los productores conocer de forma determinante a estos híbridos. Rohlf (1990) indica que la morfometría es la técnica para la descripción cuantitativa y el análisis e interpretación tanto de los morfos, como de la variación de los morfos en biología. En ese mismo orden, Leiva (2005) señala que es un método cuantitativo que se basa en la realización de mediciones de algún segmento del cuerpo, que utiliza principalmente los conceptos de forma y tamaño.
En tal sentido, el propósito de esta investigación es identificar los caracteres morfológicos externos que permitan el reconocimiento del híbrido cachamoto (Colossoma macropomum x Piaractus brachypomus) y sus parentales mediante el uso del análisis morfométrico multivariado.
II. MATERIAL Y MÉTODOS
Se analizaron 11 ejemplares de la cachama (Colossoma macropomum) macho, 11 ejemplares del morocoto (Piaractus brachypomus) hembra y 11 ejemplares del híbrido cachamoto (Colossoma macropomum x Piaractus brachypomus) con aproximadamente 600g, provenientes de estanques artificiales de la Estación Piscícola Papelón (Figura 1).
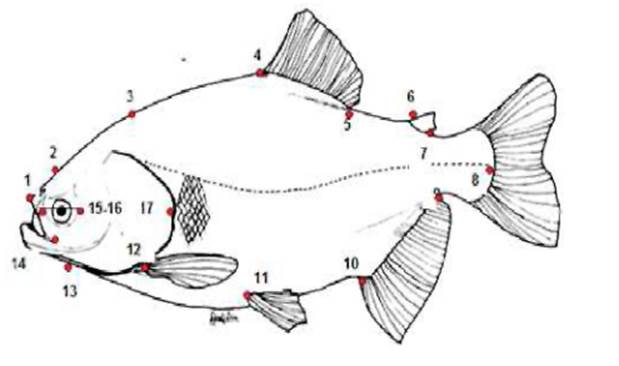
Figura 1. Localización de puntos homólogos en cachama Colossoma macropomum.
Se utilizó el método‘Truss protocol” o “cerchas” que consiste en un sistema de medidas de distancias verticales, horizontales y oblicuas entre puntos anatómicos que permiten formar entre los puntos una caja estructural, de tal manera que conformen una serie de cuadriláteros con sus respectivas diagonales; el uso de esta caja permite detectar diferencias en la forma, en todas las direcciones (Figura 2 y Tabla 1).
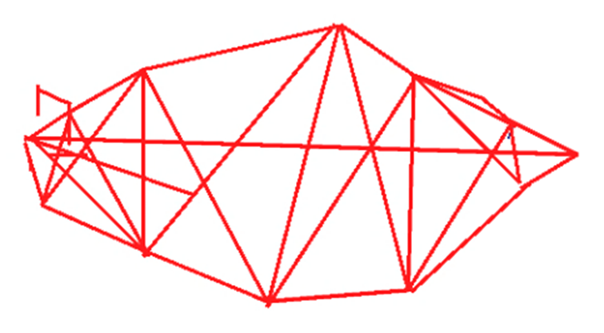
Figura 2. Protocolo de cerchas utilizado para describir la morfología externa del híbrido y sus parentales.
Tabla 1. Distancia entre puntos homólogos que indican variables medidas en el híbrido cachamoto (Colossoma macropomum X Piaractus brachypomus) y sus parentales
|
Puntos |
Distancias |
|
1 |
Largo estándar |
|
2 |
Distancia hocico a la epifiseal |
|
3 |
Distancia hocico a la aleta pectoral |
|
4 |
Distancia epifiseal a la supraoccipital |
|
5 |
Distancia epifiseal a la aleta pectoral |
|
6 |
Distancia epifiseal a la articular |
|
7 |
Distancia articular al origen aleta pectoral |
|
8 |
Distancia supraoccipital al origen aleta dorsal |
|
9 |
Distancia supraoccipital al origen aleta pélvica |
|
10 |
Distancia supraoccipital al origen aleta pectoral |
|
11 |
Distancia supraoccipital al articular |
|
12 |
Distancia aleta pectoral a la aleta pélvica |
|
13 |
Base aleta dorsal |
|
14 |
Distancia origen aleta dorsal al origen aleta anal |
|
15 |
Distancia origen aleta dorsal a la aleta pélvica |
|
16 |
Distancia origen aleta dorsal a la aleta pectoral |
|
17 |
Distancia aleta pélvica al origen aleta anal |
|
18 |
Distancia posterior aleta dorsal a la posterior aleta adiposa |
|
19 |
Distancia posterior aleta dorsal a la posterior aleta anal |
|
20 |
Distancia posterior aleta dorsal al origen aleta anal |
|
21 |
Distancia posterior aleta dorsal a la aleta pélvica |
|
22 |
Base aleta anal |
|
23 |
Distancia posterior aleta adiposa al origen aleta caudal |
|
24 |
Distancia posterior aleta adiposa a la posterior aleta anal |
|
25 |
Distancia posterior aleta adiposa al origen aleta anal |
|
26 |
Distancia posterior aleta anal a la aleta caudal |
|
27 |
Largo maxilar |
|
28 |
Diámetro ojo |
|
29 |
Largo cabeza |
|
30 |
Base aleta adiposa |
|
31 |
Ancho interorbitario |
|
32 |
Ancho pectoral (cabeza) |
Los patrones morfológicos se estudiaron mediante las siguientes técnicas estadísticas: En primera instancia, se utilizó el análisis de componentes principales (ACP) como técnica exploratoria, permitiendo resumir las direcciones principales de covariación de los datos, mediante la reducción de las variables originales a un número menor, las cuales son una combinación lineal de las variables originales y conservan toda la información contenida en estas, lo que permite una fácil interpretación. Así mismo, el análisis de componentes principales permitió estudiar aquellas variables de discriminación en términos de la morfología externa, con el fin de determinar los patrones de variación morfológica de cada una por separado. Adicionalmente los “scores” de cada ejemplar se proyectaron en cada componente principal y en el espacio canónico bidimensional en unidades estandarizadas con el fin de clasificar los ejemplares de estos peces en subgrupos homogéneos. De igual manera, se utilizó el análisis discriminante como técnica multivariante para tratar de agrupar o clasificar los individuos en grupos o subpoblaciones. Con lo cual se generaron dos funciones discriminantes canónicas que permitieron realizar pronósticos para determinar si realmente la población se ha segmentado en k grupos definidos por las variables morfológicas (Tabla 2 y Figura 3).
Tabla 2. Variables incluidas en el modelo (análisis discriminante) y estadísticas relacionadas
|
Variable |
Tolerancia |
|
Base aleta adiposa |
0 ,364 |
|
Distancia posterior aleta dorsal al origen aleta anal |
0,019 |
|
Distancia posterior aleta anal a la aleta caudal |
0 ,096 |
|
Ancho pectoral (cabeza) |
0,094 |
|
Distancia posterior aleta dorsal al posterior aleta adiposa |
0,040 |
|
Base aleta anal |
0,075 |
|
Distancia aleta pélvica al origen aleta anal |
0,083 |
|
Distancia articular al origen aleta pectoral |
0,074 |
|
Distancia posterior aleta adiposa al posterior aleta anal |
0,047 |
Figura 3. Variables que discriminan al híbrido cachamoto y sus parentales en términos de la morfología externa.
En el Tabla 3 se muestra las dos (2) funciones discriminantes y las estadísticas relacionadas que componen el modelo matemático que permite la discriminación morfológica del híbrido y sus parentales, allí se observa que la primera función explica el 79,8% de la variabilidad morfológica de las tres especies, mientras que la segunda función solo explica el 20,2%. No obstante, la correlación canónica de ambas funciones es alta (0,99 y 0,97), respectivamente. Por otro lado, el estadístico lambda de Wilks muestra los contrastes jerárquicos de las dos funciones obtenidas, en tal sentido, el primer contraste (1 ala 2), que incluye ambas funciones tomadas juntas (el modelo completo) sugiere que, el modelo permite distinguir significativamente (p<0,01) la morfología de éstas tres especies. Al mismo tiempo, el contraste para la segunda función discriminante sugiere que la misma permite discriminar significativamente (p<0,01) al menos dos de estas especies. Así pues, en la Tabla 4 se muestra la comoparación morfológica por pares de estas tres especies, donde se observan diferencias significativas (p<0,01) entre el híbrido y sus parentales, lo que verifica la capacidad de ambas funciones discriminantes para distinguir morfológicamente híbridos y sus parentales.
Tabla 3. Funciones discriminantes canónicas incluidas en el análisis y estadísticas relacionadas
|
Función discriminante canónica |
Autovalores |
% Varianza explicada |
Correlación canónica |
Contraste de las funciones (Lambda de Wilks) |
|
|
1 a la 2 |
2 |
||||
|
Función 1 |
70,33 |
79,8 |
0,99 |
0,001** |
0,053** |
|
Función 2 |
17,83 |
20,2 |
0,97 |
||
Tabla 4. Comparación morfológica por pares entre ejemplares de híbrido y sus parentales
|
Especie |
Valor del estadístico F y su significación |
|
|
Cachama negra |
Cachama blanca |
|
|
Híbrido |
51,415** |
109,360** |
|
Cachama negra |
|
162,505** |
La Tabla 5 muestra la ubicación de los centroides de la morfología de las tres especies, en cada una de las funciones discriminantes, allí se observa que la primera función distingue fundamentalmente a la cachama blanca (cuyo centroide está ubicado en la parte negativa) del híbrido y la cachama negra (cuyos centroides se encuentran ubicados en la parte positiva). Por otro lado, en la segunda función el centroide del híbrido se ubica en la parte negativa, mientras que los centroides de la cachama negra y la blanca se encuentran en la parte positiva. Dado que la primera función ha conseguido explicar el máximo de las diferencias morfológicas entre la cachama blanca y las otras dos especies (híbrido y cachama negra), es lógico que la segunda función discrimine precisamente entre el híbrido y sus parentales, que han quedado más próximas en la primera.
Tabla 5. Valor de los centroides en las funciones discriminantes
|
Especie |
Función discriminante |
|
|
Función 1 |
Función 2 |
|
|
Híbrido |
3,063 |
-5,482 |
|
Cachama negra |
7,896 |
4,077 |
|
Cachama blanca |
-10,958 |
1,405 |
En la Tabla 6 se muestran los coeficientes estandarizados de las dos funciones discriminantes, donde se observa que la primera función (la de mayor capacidad discriminativa) distingue, fundamentalmente, entre peces con una distancia posterior aleta dorsal al origen aleta anal más prolongada y una menor distancia posterior aleta dorsal a la posterior aleta adiposa. En tal sentido, dado que el único centroide negativo en esta función (Tabla 5) es el de los ejemplares de cachama blanca, se puede interpretar que, peces con una mayor prolongación de la distancia posterior aleta dorsal al origen aleta anal y una menor distancia posterior aleta dorsal a la posterior aleta adiposa tienden a ser clasificados como ejemplares de cachama blanca. Lo mismo vale decir para peces con mayor distancia base aleta caudal y menor ancho pectoral (cabeza), además de aquellos ejemplares con mayor prolongación de la distancia aleta pélvica al origen aleta anal y una menor distancia articular al origen aleta pectoral. La segunda función atribuye la mayor ponderación a la distancia posterior aleta anal al origen aleta caudal y a la distancia posterior aleta adiposa a posterior aleta anal. Puesto que ahora, el centroide del híbrido es negativo y el de la cachama negra es positivo, se puede interpretar que aquellos peces con mayor prolongación de la distancia posterior aleta adiposa a la posterior aleta anal y una menor distancia posterior aleta anal al origen aleta caudal tenderán a ser clasificados como híbridos, mientras que aquellos con una distancia posterior aleta anal al origen aleta caudal más prolongada y una menor distancia posterior aleta adiposa a posterior aleta anal tenderán a ser clasificados como cachama negra.
Tabla 6. Coeficientes estandarizados de las funciones discriminantes canónicas
|
Variable |
Coeficientes estandarizados |
|
|
Función 1 |
Función 2 |
|
|
Base aleta adiposa |
0,918 |
0,202 |
|
Distancia posterior aleta dorsal al origen aleta anal |
4,806 |
-0,817 |
|
Distancia posterior aleta anal a la aleta caudal |
-0,122 |
2,487 |
|
Ancho pectoral (cabeza) |
2,027 |
1,1568 |
|
Distancia posterior aleta dorsal al posterior aleta adiposa |
-4,487 |
-0,814 |
|
Base aleta anal |
-2,207 |
-0,546 |
|
Distancia aleta
pélvica al |
-1,922 |
0,734 |
|
Distancia articular al origen aleta pectoral |
1,940 |
-0,309 |
|
Distancia posterior aleta adiposa al posterior aleta anal |
-0,667 |
-2,486 |
III. RESULTADOS Y DISCUSIÓN
En la Tabla 7 se muestran los resultados del análisis de componentes principales sobre la morfología del híbrido cachamoto y sus parentales, donde se observa que, según el criterio de Keiser, se han seleccionado dos (2) componentes principales para el análisis , ya que para éstas, los autovalores son mayores que uno (1) y el porcentaje de varianza acumulada es de 94,29%, con lo cual podemos establecer que la dimensionalidad real del espacio morfológico (morfoespacio) en el que se encuentran los ejemplares del híbrido cachamoto (Coiossoma macropomum X Piaractus brachypomus), y sus parentales es de tamaño dos (2). Al mismo tiempo, la Figura 4 muestra una proyección de estos peces en el plano de las dos primeras componentes, donde se observa que aparentemente, existen tres (3) grupos diferenciales: híbrido, cachama blanca y cachama negra). No obstante, el caso 33, el cual corresponde a un ejemplar de cachama blanca, aparece confundido con los ejemplares del híbrido cachamoto. Lo mismo ocurre con los casos 12, 16, 17 y 19, correspondientes a ejemplares de cachama negra, los cuales muestran una ligera tendencia a agruparse o confundirse con el híbrido. Estos resultados difieren de los reportados por García y Villegas (2008) al comparar morfométricamente al híbrido cachamoto y sus parentales. Estos resultados, sugieren el uso de técnicas de análisis multivariado más refinadas, que permitan una mejor discriminación de estas especies en términos de la morfología externa, para lo cual se recomienda el uso del análisis discriminante.
Tabla 7. Autovalores de la matriz de correlación morfométrica (análisis de componentes principales) y estadísticas relacionadas
|
Componente |
Autovalores % |
Varianza explicada % |
Varianza acumulada |
|
1 |
28,74 |
89,83 |
89,83 |
|
2 |
1,42 |
4,46 |
94,29 |
En la Figura 4 se presentan los resultados iniciales del análisis discriminante , allí se puede observar que de las 32 variables morfológicas, la técnica (selección stepwise) ha seleccionado 9 variables: base aleta adiposa, distancia posterior aleta dorsal al origen aleta anal, distancia posterior aleta anal a la aleta caudal, ancho pectoral (cabeza), distancia posterior aleta dorsal al posterior aleta adiposa, base aleta anal, distancia aleta pélvica al origen aleta anal, distancia articular al origen aleta pectoral, distancia posterior aleta adiposa al posterior aleta anal, las cuales serán incluidas en la construcción del modelo matemático que permita la discriminación del híbrido cachamoto y sus parentales en términos de su morfología externa.
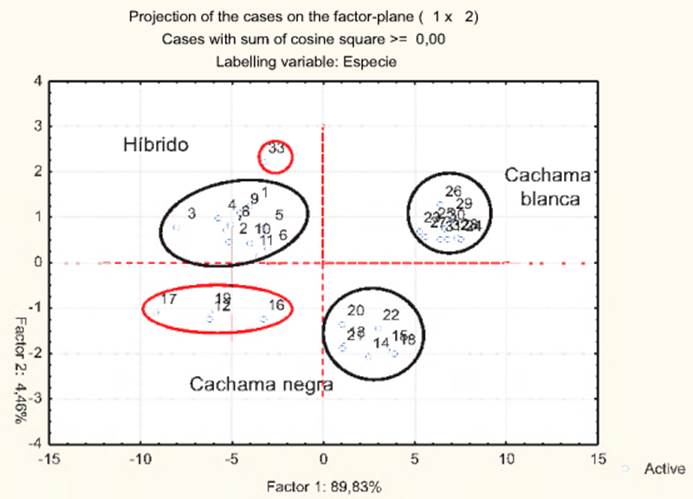
Figura 4. Proyección de los scores morfológicos para ejemplares de híbrido cachamoto y sus parentales en el plano (CP1*CP2).
Lo mismo ocurre con los ejemplares de carachama negra (casos 12, 16, 17 y 19), los cuales en principio (ver proyección de los scores morfológicos en el plano CP1*CP2), mostraron una ligera tendencia a confundirse con los híbridos, no obstante, al revisar las puntuaciones discriminantes de estos peces en la segunda función (4,433; 3,495; 3,255; y 3,672), respectivamente, se observa que los mismos fueron clasificados correctamente en el grupo al que pertenecen (carachama negra). De igual forma, esto se evidencia en las probabilidades condicionales P(D > d/ G = g) de estos peces (0,730; 0,844; 0,741 y 0,723), respectivamente, lo que sugiere que se trata de peces centrados en el grupo den el que han sido clasificados (carachama negra).
La Tabla 8 muestra los resultados de la clasificación morfológica del híbrido y sus parentales, allí se observa que para el caso 33 (ejemplar de cachama blanca), el cual fue confundido con un híbrido cuando en el análisis de componentes principales, en este caso los resultados sugieren que este ejemplar se corresponde con uno de cachama blanca, y no así con un híbrido como lo sugirió el ACP. Al observar la puntuación discriminante de este individuo en la primera función discriminante (-11,398), esto hace que sea clasificado correctamente en el grupo al que pertenece (cachama blanca). Así mismo, la probabilidad condicional P(D > d/ G = g) de este ejemplar tiene un valor alto (0,723), lo que permite afirmar que se trata de un pez bastante centrado en el grupo en el que ha sido clasificado.
Tabla 8. Clasificación morfológica del híbrido cachamoto y sus parentales
|
|
|
Número de caso |
||||
|
|
|
12 |
16 |
17 |
19 |
33 |
|
Grupo real |
|
|
Cachama negra |
|
Cachama blanca |
|
|
|
Grupo pronosticado |
|
Cachama negra |
|
|
|
|
|
P(D > d/ G = g) p |
0,730 |
0,844 |
0,741 |
0,723 |
0,723 |
|
|
gl |
2 |
2 |
2 |
2 |
2 |
|
Grupo mayor |
P(G = g/ D = d) |
1,000 |
1,000 |
1,000 |
1,000 |
1,000 |
|
|
Distancia de Mahalanobis al cuadrado hasta el centroide |
0,628 |
1,210 |
1,630 |
0,648 |
0,648 |
|
|
Grupo |
Híbrido |
||||
|
|
P(G = g/ D = d |
0,000 |
0,000 |
0,000 |
0,000 |
0,000 |
|
Segundo grupo mayor |
Distancia de Mahalanobis al cuadrado hasta el centroide |
129,01 |
113,80 |
110,13 |
114,30 |
147,73 |
|
Puntuaciones discriminantes |
Función 1 |
8,604 |
8,833 |
8,876 |
8,8591 |
-11,398 |
|
Función 2 |
4,433 |
3,495 |
3,255 |
3,672 |
0,731 |
|
La Figura 5 muestra una dispersión de las tres especies sobre el plano definido por las dos funciones discriminantes, allí se observa que la primera función (eje horizontal), permite distinguir fácilmente las cachamas blancas de los híbridos y cachamas negras. De igual forma, la segunda función (eje vertical), permite diferenciar fácilmente al híbrido de la cachama negra.

Figura 5. Diagrama de dispersión de los tres grupos (híbrido, cachama negra y cachama blanca) en las dos funciones discriminantes canónicas.
IV. CONCLUSIONES
El análisis de componentes principales no mostró una clara diferenciación morfológica entre el híbrido y sus parentales, lo que evidencia una desventaja de esta técnica multivariada. El análisis discriminante logró reducir el total de variables morfológicas (32) a nueve (9), las cuales permitieron construir un modelo matemático para lograr una mejor discriminación morfológica entre el híbrido y sus parentales.
Se construyeron dos funciones discriminantes canónicas con una alta capacidad para distinguir morfológicamente al híbrido de sus parentales. La primera función logró discriminar a la cachama blanca del híbrido y cachama negra, mientras que la segunda función mostró capacidad para distinguir al híbrido de sus parentales.
Los resultados del análisis discriminante sugieren que, peces con una mayor prolongación de la distancia posterior aleta dorsal al origen aleta anal y una menor distancia posterior aleta dorsal a la posterior aleta adiposa, además de aquellos con mayor distancia base aleta caudal y menor ancho pectoral (cabeza), así como aquellos ejemplares con mayor prolongación de la distancia aleta pélvica al origen aleta anal y una menor distancia articular al origen aleta pectoral tienden a ser clasificados como ejemplares de cachama blanca. Por otro lado, se puede inferir que aquellos peces con mayor prolongación de la distancia posterior aleta adiposa a posterior aleta anal y una menor distancia posterior aleta anal al origen aleta caudal tenderán a ser clasificados como híbridos, mientras que aquellos con una distancia posterior aleta anal al origen aleta caudal más prolongada y una menor distancia posterior aleta adiposa a la posterior aleta anal tenderán a ser clasificados como cachama negra. El uso del análisis discriminante permitió una clasificación correcta de cada especie, descartando cualquier clase de confusión morfológica entre el híbrido y sus parentales.
No obstante, los resultados de este trabajo, no son suficientes para asegurar que existen diferencias morfológicas entre el híbrido cachamoto y sus parentales, aun cuando éstas se evidencian en el análisis estadístico multivariado (discriminante) aplicado a ejemplares de híbrido cachamoto y sus parentales, puesto que existe una variabilidad en la talla y peso de los mismos, por lo que el factor alometría pudiera estar afectando considerablemente los resultados de esta investigación.
V. REFERENCIAS BIBLIOGRÁFICAS
Campos, M. 2007. Los recursos zoogenéticos de Venezuela. Caracas (Venezuela): Ministerio del Poder Popular para el Ambiente.
García T. y Villegas D. 2008. Análisis morfométrico comparado del híbrido cachamoto (Colossoma macropomum x Piaractus brachypomum) y sus parentales, cachama (Colossoma macropomum) y morocoto (Piaractus brachypomum). Informe de Investigación. Guanare (Venezuela)
Heredia, B. y González, J. 1986. “Estudios preliminares del crecimiento del híbrido de cachama (Colossoma macropomum) y morocoto (Piaractus brachypomum) alimentados con dietas artificiales.” En XXXVI Convención Anual de la ASOVAC. Valencia (Venezuela).
Leiva, M., J. I. Mojica, R. Royero. 2005. “Revisión taxonómica del género Pimelodella, Eigenmann y Eigenmann,1888 (pisces, siluriformes: heptapteridae) de la región transandina de Colombia.” Acta Biológica Colombiana 10 (1): 95
Orozco, J. J. 1990. Estudio de crecimiento y de producción de cachama negra (Colossoma macropomum) y la cachama blanca (Piaractus brachypomum) a densidades altas en tanques y jaulas flotantes. Informe de investigación. Lieja (Bélgica).
Rohlf, F. J. 1990. “Morphometrics.” Annual Review of Ecology and Systematics 21: 299-316.